There is a double-standard in psychology that claims that humans have unique socio-cognitive skills, and a prominent explanation for this is the Social Intelligence or Social Brain Hypothesis: that human brains are outsized due to the enormous complexity of our social relations. I have noted in previous posts that the skills psychologists assume are human-specific have been found in other animals, casting doubt on whether these social abilities are as unique as they insist, but we also have reason to doubt whether these social abilities are related to having a large brain.
This should be immediately obvious from the fact that human beings, who most researchers (implicitly or explicitly) deem the most intelligent, socially or otherwise, have a brain 5 times lighter than the brain of the average sperm whale (Marino, 2004). This is not altogether surprising as the sperm whale is also one of the most massive creatures (ever) on earth, and there is a positive relationship between body size and brain size (Roth and Dicke, 2005; see figure 1).
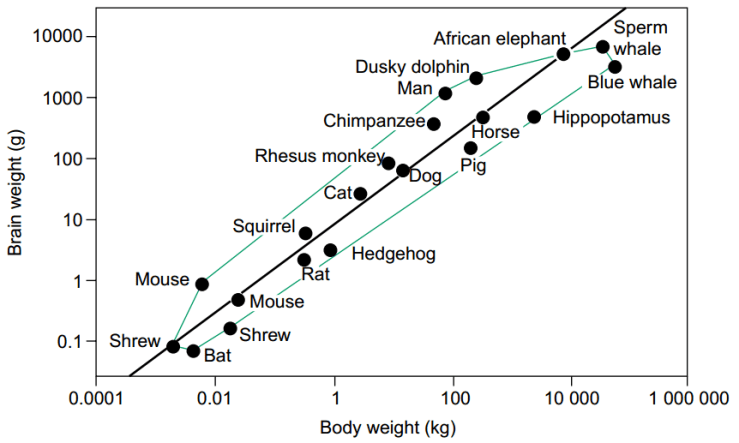
Figure 1: log of body weight against log of brain weight for 20 mammals. The black line represents the regression equation for this set; the green line connects the farthest deviations away from this regression. From Roth and Dicke (ibid)
However, taking body weight into account does humans and similarly “intelligent” creatures no favours either, as brains increase absolutely as body size increases, but decrease as a proportion of body size as body size decreases (since neural cells can’t get too small before they lose functionality). This is why mice have brains that take up around 10% of their mass (ibid; see figure 2) but are not currently thought to be 5 times more intelligent than humans. So far we can see that measuring brain size in either absolute or relative terms does not appear to grant us the hierarchy we expect to see.

Fig 2: log body weight plotted against log relative brain weight for a selection of 20 mammals. From Roth and Dicke (ibid)
However, as figure 1 demonstrates, humans are an outlier on the graph (a regression of this graph reveals a predicted relative human brain size of 0.3% of body weight, 7 times less than reality). So another measure of brain size examines how far a species deviates from an expected value based on a related taxon, known as “encephalisation quotient” or “EQ”. An EQ of 1 would represent matching the predicted brain weight according to their body weight. Humans under this formula have an average EQ of 7.4-7.8, indicating a brain over 7 times larger than expected for the taxon, and the highest EQ of all. The formula for deriving this value depends particularly on how we calculate the regression line from which we compare species: comparing humans amongst primates puts the constant at .28 (Williams, 2002), but for comparing amongst mammals and birds, it could be between .76 (Martin, 1981) and .56 (Kuhlenbeck, 1973).
The abstractions made by the EQ formula renders it sensitive to which species are included in the sample, but also in how one measures mass. Overall weight is an indirect way of measuring mass, but this elides the differences in cell types both in the brain (e.g. white matter and grey matter) and in the body (e.g. muscle and adipose tissue; see Schoenemann, 2004). Readers may be familiar with body mass index or BMI, and how such a measure can fail to distinguish the difference between, for instance, a heavyweight boxer and a sumo wrestler: a similar principle means that nimble capuchin monkies have a greater EQ than gorillas (Marino, 1998; Jerison, 1973; Gibson et al., 2001), despite capuchins performing worse in cognitive tests (Deaner, et al, 2007). For this reason, Herculano-Houzel et al (2007) were able to reinterpret this finding, suggesting that great apes like gorillas might be disadvantaged by EQ by having a body relatively larger than one would expect, given their brain size.
If we take a more cell or structure-based approach to brain volume, we are confronted with similar problems. If we look at the average number of neurons or synapses,we can see that the species with the most cortical neurons and synapses is the African elephant, not any primate (Roth, Stamenov, and Gallese, 2002). The vaunted human cerebral cortex, the folded outer layer of the brain, is only as large as one would expect for a primate with our body size, and holds around the same proportion of neurons as in other mammals’ cerebral cortex, 19% (Herculano-Houzel, 2009). However, given that corvids famously do very well in cognitive tests with a radically different (and considerably smaller) cortical structure, perhaps we should look beyond the brain when assessing intelligence.
Organisms are products partly of their environment, and their physiology and behaviour will reflect this difference. Delphinids and corvids are often investigated for their complex social behaviour that may have some resemblances to human social intelligence (Xitco, Gory, and Kuczaj, 2001; Emery, 2005). However, these animals are not merely differently shaped primates: they have unique brain structures that reflects the different environments that they have evolved within, and to compare them directly to human beings is a mistake. For example, dolphins have excellent hearing but poor vision in comparison to primates which reflects their usage of ultrasonic communication and perception (Barrett and Würsig, 2013). Behavioural observations by Brownlee and Norris (1994) have suggested that dolphins can use echolocation to pick up on emotional cues, and identify which of their conspecifics is full and which has not eaten by echo locating their stomach, which is something Barrett and Würsig (op. cit.) point out as having no human analogue. How can we apply our standards of social intelligence, such as deception, to an animal capable of using sonic perception to “see” its’ group member’s empty stomach?
The atheoretical search for a measurement of brain size barely conceals a search for a measurement that confirms human suspicions of our cognitive superiority in all regards. One yardstick after another is discarded when it does not grant our species pole position: but it is the theory that all brains, regardless of species, should fit linearly along a scale of intelligence according to sizethat should be done away with, especially given the lack of success it has seen thus far. Ultimately, the idea of intelligence as a distinct entity, located in the brain, reducable to a single linear scale from idiots to savants, is the residue of race science in psychology, no longer used to prop up white supremacy (at least not directly) but still shaping cognitive science.

In contrast, Barrett, Henzi, and Rendall, (2007) and others have shown that complex behaviour results not merely from cognition and brain mass, but from the interaction of the organism with its environment over time. For example, Strum, Forster, and Hutchins, (1997) offered a reinterpretation of apparently “Machiavellian” male baboons (E.Q between 1.74 and 2.05), who utilise a tactic they call “sleeping near the enemy” to displace older males from females. Older males could resist attacks from younger rivals during daytime, when the troop was on the plains, but at night, the troop would retire to the cliffs to sleep. Younger males would sleep closer to the males and attack them at night. Strum and authors proposed that instead of seeing this as evidence of the younger baboons collecting and reflecting on this information, formulating plans, imagining the future, etc., it was a result of the environment being unable to afford the older male help from allies or an escape. The cliffs are narrow and have limited pathways in comparison to the plains, and this environment forces a conflict just between the two, on the young baboon’s terms. This is not to say the baboons do not use their brains, or that the environment is entirely responsible for this behaviour: it is evidence that the environment places constraints or offers advantages for certain behaviours to exploit, and that re-interpretations that use this simplified, ecological, and dynamic view of complex behaviour are possible.
